NB: The
information at this web site was published in the following paper:
Olsen, P. E. and Huber, P., 1998, The oldest Late Triassic footprint
assemblage from North America (Pekin Formation, Deep River basin, North
Carolina, USA). Southestern Geology, v. 38, no. 2, p. 77-90.
The oldest Late Triassic footprint assemblage from North
America (Pekin Formation, Deep River Basin, North Carolina, USA)
Paul E. Olsen, Lamont-Doherty Earth Observatory of Columbia University,
Rt. 9 W, Palisades, NY 10964, email polsen@ldeo.columbia.edu
and Phillip Huber, Department of Education, University of Bridgeport, Bridgeport,
CT 06601
ABSTRACT
An assemblage of reptile footprints from the abandoned Pomona Terra-cotta
and an adjacent active quarry in the middle Pekin Formation of the Sanford
subbasin of the Deep River basin is the oldest track faunule recognized
to date in strata of Late Triassic age in Eastern North America. The most
common taxon is a possibly new genus of pentadactyl ichnite similar to,
but distinct from, Brachychirotherium. It may lack manus impressions,
has a strong tendency to be functionally tridactyl, and has an extremely
shallow digit V impression. At least one track is over 30 cm in length.
Also present is the quadrupedal, probably phytosaurian ichnite Apatopus
lineatus, based on a trackway, and several small (<15 cm) bipedal
and tridactyl forms that are probably dinosaurian. Other more poorly preserved
forms are present. Apart from Apatopus, none of the tracks fit into
recognized Newark ichnotaxa. The age of this track assemblage is early
Tuvalian (early Late Carnian of the Late Triassic), based on associated
tetrapod skeletal and macro- and micro-floral remains. This middle Pekin
footprint assemblage is thus distinctly different, and older than. all
other Newark Supergroup footprint assemblages. It is important because
it represents a transitional stage between the well known Middle Triassic
assemblages and the more typically Newarkian Late Triassic assemblages.
The apparently dinosaurian ichnites from this horizon are therefore arguably
among the oldest in the world.
INTRODUCTION
Footprints comprise by far the most common evidence of tetrapods in
the Newark Supergroup of eastern North America. They have been the subject
of fairly continuous research since the first dinosaur footprint was described
by Edward Hitchcock in 1836 (Hitchcock, 1836; Olsen et al., 1997), and
they provide a rich source of biostratigraphic, paleoecological, behavioral,
and physiological information (e.g. Olsen and Galton, 1977; Olsen, 1988;
Lockley, 1991; Farlow, 1981; Farlow and Chapman, 1997). Despite the over
160 years of study devoted to Newark Supergroup tetrapod footprints, the
assemblages from the Early Jurassic age strata have received most of the
attention, with antecedent 30 million years of Newark track assemblages
receiving relatively short shrift. Indeed, it is within only the last 45
years that the Triassic-age assemblages have been recognized as distinct
in composition (Baird, 1957; Olsen and Baird, 1986; Fraser and Olsen, 1996)
with most of the occurrences still being known from superficial descriptions
(e.g. Olsen. 1988; Olsen et al., 1989). Here we describe the oldest known
Late Triassic age footprint assemblage in the Newark Supergroup, that from
the middle Pekin Formation of the Deep River Basin of North Carolina.
GEOLOGICAL PROVENANCE
The Deep River basin of North and South Carolina is the southernmost
exposed of a extensive series of rift basins formed during the Triassic
and Early Jurassic break up of Pangea (Fig. 1). The footprint assemblage
described herein, comes from the Pekin Formation of the northern part of
the Sanford subbasin of the Deep River Basin, which is the oldest formation
recognized in the basin. Presently, the Pekin Formation is placed within
the Chatham Group of the Newark Supergroup (Weems and Olsen, 1997) and
comprises tectonostratigraphic sequence II (TS II), the oldest, widespread
rift sequence in the Central Atlantic Margin rifts (Olsen, 1997).
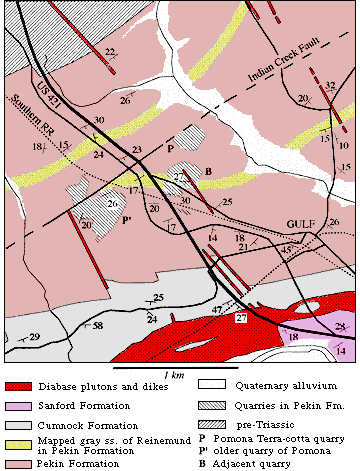
| All of the footprints described here come from two quarries
called the Pomona Terra-cotta Co. quarry (now abandoned) and an adjacent
quarry developed in the lower part of the middle Pekin Formation (Fig.
2). As described by Reinemund (1955), the Pekin Formation in the northern
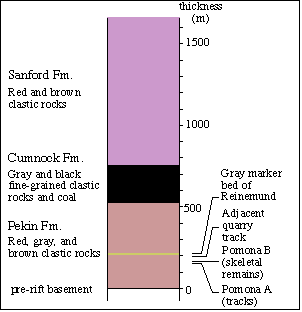
Sanford subbasin, in the vicinity of these quarries, is about 530 - 550
m thick (Fig. 3). Its basal beds tend to consist of about 90 m of gray
or brown conglomerate and sandstone followed by mostly lenticular beds
of red, brown or purple claystone, siltstone sandstone, and locally conglomeratic
arkosic sandstone. A gray sandstone and siltstone sequence is exposed near
the top of the section in the main part of the adjacent quarry and this
interval is a marker bed mapped throughout the northern Sanford subbasin
by Reinemund (1955) (Fig. 2). Its position, according to Reinemund, is
about 210 m above the base of the section. It is this unit that produced
the well known plant assemblages described by Hope and Patterson (1969),
Delevoryas and Hope, (1975), and Axsmith et al., (1995), among others.
All of the footprints occur below this bed, probably within about 100 m.
Most of the footprints from Pomona A, however were found in rubble.
The important assemblage of tetrapod bones described by Baird and Patterson
(1968) and Huber et al. (1993) (Table 2) apparently come from the southern
side of the Pomona quarry, here termed Pomona B. During the 1970's Pomona
B was being filled in and the existing exposures did not allow for the
section to be measured in detail. One track was found in 1989 in rubble
from the adjacent Pit. |
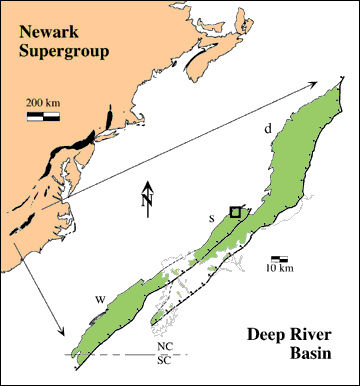 |
Figure 1: Deep River basin of North and South Carolina and
Newark
Supergroup of eastern North America. Box shows position of
footprint localities (see Fig. 2): d, Durham subbasin; s,
Sanford
subbasin; w, Wadesboro subbasin. Gray line shows limit of
Coastal
Plain. |
 |
The projected trace of the Indian Creek fault shown by
Reinemund (1955) should pass through the Pomona quarry, and it should have
between 100 and 250 m of normal displacement. However, a fault of this
magnitude was not observed by PEO in either the Pomona or adjacent quarry
over 15 year of visits, and hence the fault presumably passes to the immediate
east of the Pomona quarry, as it is shown in Figure 2. However, several
small faults were seen in the Pomona and adjacent quarries. From their
narrow gouge zones, the similarity of facies on both sides of the faults,
and the lack of major repetition of strata, these faults probably have
displacements of a few meters. Unfortunately, one of these small faults
separates the Pomona A section from the Pomona B section, and this and
the other small faults make it impossible to compile a complete section
from the plant-bearing gray sandstone marker bed in the adjacent quarry
through the main footprint-bearing units in the Pomona quarry, given the
exposures present in the 1970's.
The tabular geometry of the footprint bearing unit in Pomona A and the
presence of conchostracans in the plant-bearing strata of the adjacent
quarry (Olsen et al, 1989) suggests some ponded water. In combination with
the presence of tilted beds of ripple-cross laminated sandstone and lenticular
cross-bedded sandstones, the environment of deposition was probably shallow
lacustrine, paludal, marginal lacustrine, and fluvial during the deposition
of the units exposed in the Pomona and adjacent quarries. The absence of
caliche and evaporites and the intense bioturbation and abundant plant
foliage (even in red units) suggests persistently humid conditions. |
Figure 2: Map of geology in vicinity of middle Pekin
Formation footprint localities. Maps are based on digitally
superposed images of the U.S.G.S. 7.5 minute Goldston
Quadrangles and maps of Reinemund (1955; plate 1-central,
and plate 4). |
|
 |
 |
|
Figure 3: Section of Triassic rocks in northern
part
of Sanford subbasin: based on Reinemund (1955). |
Figure 4: Exposure of footprint-
bearing strata at north end of
Pomona A. White bar is 1 m and is
positioned just below main
footprint-producing unit (Table 1). |
|
Table 1 Measured section at footprint-producing area on the north
side of the Pomona Terra-cotta quarry. Section measured December
29, 1977. * denotes main track-bearing unit.
| thickness |
lithological
description |
fossils, other |
| normal fault |
|
|
| +1 m |
red hackly mudstone |
?Scoyenia |
| 0.5 m |
purple-brown,
green up hard siltstone |
roots |
| 0.9 m |
deep purple,
light purple up mudstone |
hematitic nodules |
| 0.4 m |
green-purple
hard siltstone and fine, sandstone |
footprints, roots,
Scoyenia |
| 3.0 m |
red fissile siltstone |
roots |
| 1.0 m* |
greenish-purple
and red, hard, flaggy siltstone and fine sandstone |
footprints, plant
foliage, roots, Scoyenia |
| 1.8 m |
red massive
mudstone, fissile upward |
fish scales,
coprolites, reptile teeth, plant foliage, Scoyenia |
| 0.2
m |
green and red,
hard, massive mudstone |
|
| 1.9
m |
red, greenish up, faintly laminated conchoidally
fracturing, siltstone |
| +1m |
green and
red poorly bedded mudstone |
roots, casts
of in situ plant stems |
| covered |
|
|
| FOOTPRINT ASSEMBLAGE
Tracks from Pomona A include Apatopus lineatus, abundant cf.
Brachychirotherium
spp., small three-toed dinosaurian tracks, and several unidentified forms.
Most of the tracks could not be recovered and have been destroyed. A trackway
consisting of a natural cast of three successive manus-pes set on a large
transported block demonstrates the presence of
Apatopus lineatus
Baird 1957 (Fig. 5). According to Baird (1957),
Apatopus lineatus
is diagnosed as a quadrupedal ichnite with a long, narrow pentadactyl pes
with the digits in increasing length V, I, II, III, IV, and a short manus
symmetrical around digit III. Usually only digits I, II, and III of the
pes impress distinctly, and this is the case with the Pekin trackway (Figs.
6, 7). Individual manus-pes sets (Fig. 6 C-E) are indistinguishable from
the type material described by Baird (1957, Plate 3, Fig. 1). |
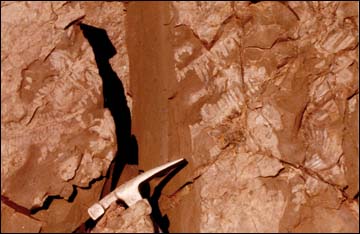 |
Figure 5: Plant foliage (mostly cycadeoid) preserved as red
clay films in red mudstone from within the main track bearing-
unit (Table 1). |
|
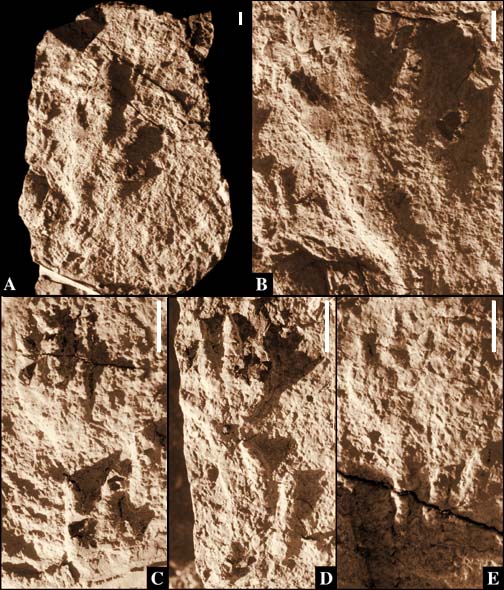 |
Figure 6: Large slab with natural
casts of Apatopus lineatus and very large
brachychirothere-like possible
example of new genus 1; slab not collected.
A, photograph of entire slab
digitally corrected for parallax; B, possible
example of new genus 1; C-D,
successive manus-pes sets of Apatopus lineatus.
Scale for all is 5 cm. |
In addition, Baird's (1957, Fig. 8) figured trackway, based on dissociated
blocks, is matched closely by the Pomona trackway (Fig. 7), which is confirmation
of his reconstruction.Baird (1957) assigned Apatopus lineatus to
the Phytosauria because of general similarities between the trackway of
Apatopus and living crocodilians, the correspondence between the reconstructed
skeleton of Apatopus and the reconstructed pes and manus, and the
overlapping stratigraphic ranges of phytosaurs. Parrish (1986) has questioned
this assignment on functional grounds, but has produced a forward model
of a phytosaur track based on known osteology (his Fig. 4.9) that is as
close to Apatopus as can be expected, given the limitations of the
method. We argue that a functional argument is inherently weaker than one
based on anatomical similarity, and therefore concur with Baird's (1957)
original assignment.
Figure 7: Outline drawing of trackway of Apatopus lineatus
(A) compared to type trackway as reconstructed by Baird 1957 (B). B is
redrawn from Baird (1957). |
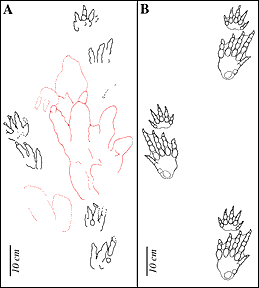 |
Figure 8: Slabs of footprints from Pomona A: A, new genus
1 and vague undetermined
tracks (slab in collection of James L. Mashburn of Sanford,
North Carolina); B, slab
with natural casts of trackway of undetermined ?brachychirothere
with possible manus
(not collected); C, large slab with deep but sloppy trackway
of new genus 1 (not
collected); D, natural casts of partial ?brachychirothere
and good dinosaurian pes
(not collected). Scale for all is 10 cm. |
 |
 |
The most abundant footprints from Pomona A are somewhat
similar to
Brachychirotherium in having a pentadactyl pes with the
digit III being longest, and digit V being very reduced (e.g. Haubold,
1971) (Figs. 8, 9). However, unlike
Brachychirotherium, the impression
of digit V is very weak and far posterior of its normal position (Fig 8),
and digit I is small with the pes beingunctionally nearly tridactyl. In
addition, none of the specimens from the Pekin Formation have an unequivocal
manus impression. In the nearly tridactyl form of the pes and absence of
a manus impression, the Pomona A forms resembles Parachirotherium postchirotheroides
of Kuhn (1958) from the Gipskeuper (Early Carnian age) of Bayreuth, in
Germany. As figured by Kuhn (and Haubold, 1971; 1986), however, digit V
is too far anterior. As exemplified by the clearest example (Figs. 8A,
9A) the track is different than any described form and probably should
be named a new genus, which we call new genus 1 for this paper. However,
we cannot name a new taxon at present, because, none of the specimens or
casts of new genus 1 reside in institutions. |
Figure 9: New genus 1: A, detail from slab in
Fig. 8A; B,
detail of lower track shown in Fig. 8C. Scale
is 5 cm. |
|
The absence of clear pads precludes a detailed osteological reconstruction
of the pes of new genus 1. However, it is clear that the track maker had
a reduced digit V, a short digit I, and a long metatarsal axis. The pes
skeleton of Postosuchus kirkpatricki (Chatterjee, 1985) is comparable
to new genus 1, and it is interesting that Chatterjee has reconstructed
Postosuchus
as bipedal, which agrees with the new genus. In addition, the probably
rauisuchian teeth from Pomona B could be a postosuchid. However, Postosuchus
has a temporal range (Carnian and Norian) much greater than new genus 1
would seem to have. More track material with better defined pads is needed
for rigorous analysis, however.
On the same slab as the Apatopus trackway (Fig. 6A, B) is a natural
cast of a very large (~35 cm) pentadactyl pes. Again, there is no manus
impression, but this form differs from new genus 1 in having a proportionally
longer digit I, shorter digit III, and more anterior and distinct digit
V. Superficially the pes is comparable to large chirotheriids and brachychirotheriids
(e.g. Haubold. 1971). It is possible, however, that the differences between
this very large form and new genus 1 could be due to ontogenetic allometric
changes similar to those seen in Early Jurassic theropod dinosaur tracks
(Olsen et al., 1997). Without specimens of intermediate size, it is impossible
to test this hypothesis.
| Tridactyl ichnites, represented by one trackway, are present
at Pomona A and the adjacent quarries (Fig 8D, 9). Two of these have vague
possible manus impressions, but without trackway confirmation, the association
could be fortuitous. All the tridactyl forms have a widely splayed pes,
with proportions and size similar to the Jurassic ichnogenus
Anomoepus
(see Lull, 1953). However, all of the Pekin tridactyl ichnites have the
characteristic placement of the metatarsal phalangeal pad of digit IV directly
in line with the axis of digit III. There is no sign of the distinctive
pentadactyl manus of Anomoepus as well. The three-toed pes and bipedal
trackway may be shared-derived characters of the Dinosauria. Because none
of the Pekin tridactyl forms have distinct phalangeal pads we do not attempt
further analysis, except to note that they are plausibly, but not definitively
dinosaurian.
There are a number of other traces from Pomona A that indicate the presence
of other ichnotaxa (e.g. Fig 8D, left). However, these are all to poor
to warrant detailed description. They indicate significant additional diversity
in this assemblage. |
 |
Figure 10: Dinosaurian tracks from Pomona A and the adjacent quarry:
A,
detail of track shown in Fig. 8D, right; B, pes and possible manus
(Yale
Peabody Museum 55875) ; C, natural cast of right pes from the adjacent
quarry (incorrectly noted as not collected in Olsen, 1998); D, part
of large
slab of vague natural casts of dinosaurian pedes (not collected).
Scale is 5
cm. |
|
ASSOCIATED FAUNAL AND FLORAL REMAINS AND AGE
Pomona B has produced an important although fragmentary tetrapod skeletal
assemblage (Baird and Patterson, 1968). Based on the small fault offset
between Pomona A and Pomona B, the footprint assemblage is no more than
40 m below the unit that produce the tetrapod assemblage, and probably
significantly less. Most distinctive is the rotund dicynodont synapsid
Placerias
cf. P. hesternus and aetosaur scutes assignable to Longosuchus
meadi. According to J. L. Mashburn and D. Baird (1974, pers. comm.)
the Placerias, at least, were originally articulated specimens prior
to blasting. Based on a correlation web with the Chinle group and the European
section, these indicate an early Tuvalian (early Late Carnian) age (Huber
et al., 1993). Also present are indeterminate phytosaur material and indeterminate
probable rauisuchian teeth. This assemblage comprises the type for the
Sanfordian land vertebrate faunachron of Huber et al. (1993), which correlates
to the Otischalkian land vertebrate faunachron of the Chinle Group of the
western United States.
Table 2:Vertebrate osteological remains and tracks (*)
from the Pomona Terra-cotta and adjacent quarries.
| Taxon |
Reference |
Horizon |
| Synapsida |
|
|
| Kannemeyeriidae |
|
|
|
Placerias cf. P. hesternus |
Huber et al., 1993 |
Pomona A |
| Reptilia |
|
|
| Archosauria |
|
|
|
Phytosauria |
|
|
|
Phytosauria indet. |
Baird & Patterson, 1968 |
Pomona A, Pomona B, adjacent quarry |
|
*Apatopus lineatus |
Olsen and Huber, 1998 |
Pomona B |
|
Suchia |
|
|
|
Longosuchus cf. L. meadei |
Huber et al., 1993 |
Pomona A |
| Rauisuchia indet. |
Huber et al., 1993 |
Pomona A |
| ?Suchia |
|
|
| *cf. Brachychirotherium
sp. |
Olsen and Huber, 1998 |
Pomona B |
| ?Dinosauria |
|
|
| *undetermined |
Olsen and Huber, 1998 |
Pomona A, Pomona B |
A palynoflorule from the basal Pekin Formation of the Sanford basin
(Cornet , 1977) and the Boren quarry has produced an extensive floral assemblage
(Hope and Patterson, 1969; Delevoryas and Hope, 1975; Olsen et al., 1989;
Axsmith et al., 1995) suggest correlation of the lower to middle Pekin
Formation with tectonostratigraphic sequence II (TS II, Olsen, 1997) of
the Richmond and Taylorsville basins. The vertebrate assemblage also suggests
a correlation with TS II of the Fundy basin of the Canadian Maritimes and
the Argana Basin of Morocco (i.e. Timezgadiwine Formation).
COMPARISONS TO OTHER ASSEMBLAGES
There are two accounts of footprints from the Newark Supergroup that
are probably older than the Middle Pekin assemblage. The oldest is from
the Honeycomb Point Formation of the Fundy basin of New Brunswick, Canada
(Olsen, 1997), but these ichnites are very poor and have never been described.
Shaler and Woodworth (1899) figure outline drawing of footprints from the
"Productive Coal Measures" of the Richmond basin. These strata belong to
the lower part of TS II and are probably older than the footprint assemblage
from the middle Pekin. The drawings are, however, inadequate for comparison
to other tracks and the whereabouts of the specimens is unknown.
Abundant and diverse footprint material has been recovered from tectonostratigraphic
sequence III (TS III) in many other Newark Supergroup basins. Nonetheless,
the only undoubted member to the Pekin footprint assemblage shared by younger
Newark Supergroup faunules is Apatopus lineatus. The oldest assemblage
that has been described in TS III is that from the upper Stockton and lower
Lockatong Formation of the Newark basin of New York, New Jersey, and Pennsylvania
(Olsen and Flynn, 1989; Olsen, 1988; Baird and Olsen, 1986) and the roughly
coeval tracks from the Cow Branch Formation of the Dan River basin of North
Carolina and Virginia (Olsen et al., 1978; Baird and Olsen, 1986; Fraser
and Olsen, 1996). All of these are Late Carnian (late Tuvalian) in age
and are associated with tetrapod skeletal taxa of the Conewagian land vertebrate
faunachron (Huber et al., 1993). The Late Carnian age assemblages are basically
the same as early and middle Norian assemblages (also in TS III) that are
known from many localities and many Newark Supergroup basins (Baird, 1954;
Baird, 1957; Olsen and Baird, 1986; Olsen, 1988; Olsen et al; 1989). These
are dominated by Brachychirotherium, Apatopus, Rhynchosauroides
and dinosaurian taxa, notably Atreipus and Grallator, and
are associated with skeletal forms of Neshanician and lower Cliftonian
land vertebrate faunachrons (Huber et al., 1993).
Otischalkian strata of the Chile Group have produced very little footprint
material (Lucas and Huber, 1997) and the younger Chinle assemblages have
nothing in common with the Pekin track faunule. A greater diversity of
footprints is known from the European lower Keuper (km1-km3), notably with
the lower Gipkeuper (km2) that produced Parachirotherium postchirotheroides.
The Argana and Ourika basins of Morocco have also produced a poorly
known assemblage of footprints, which at first glance appears potentially
similar to that from Pomona A (Biron and Dutuit, 1981). Apatopus lineatus
appears to be present in the Ourika basin (Biron and Dutuit, 1981) along
with a tetra-or pentadactyl tracks (Quadridigitatus dubius of Biron)
and a few unnamed tridactyl forms. Biron named two tri- tetra-or pentadactyl
tracks, Tridactylus manchouensis and Anomoepus moghrebensis,
that were recovered from the Timezgadiwine Formation of the Argana basin
(comprising TS II of Olsen, 1997). These could be poorly preserved examples
of new genus 1, however restudy and collection of new material is clearly
needed. The age of the Timezgadiwine Formation is early Tuvalian (Late
Carnian) and that of the tracks in the Ourika basin is constrained only
to Carnian on the basis of pollen and spore assemblages (Cousminer and
Manspeizer, 1976).
IMPORTANCE OF THE PEKIN ASSEMBLAGE
The footprint assemblage from the Pekin Formation of the Deep River
basin is the oldest in eastern North America that is based on material
good enough to analyze. Coming from strata of early Tuvalian age, it is
the same age as the oldest known dinosaurs (Lucas and Long, 1992; Lucas
and Huber, 1997). King and Benton (1996), suggest that all published records
of dinosaur footprints of Middle Triassic age are doubtful, and therefore
the tracks of dinosaurian aspect of the Pekin assemblage are among the
oldest in the world. That said, it is also true that since both saurischians
and ornithischians are present in late Tuvalian age strata, early Tuvalian,
or even late Middle Triassic age dinosaurs plausibly existed. In addition,
the feet of the earliest known dinosaurs, such as Herrerasaurus
retain very primitive proportions with both long digits I and IV (Novas,
1993). Therefore, it is likely that it may be quite difficult to recognize
the pes of very early dinosaurs because they might resemble a brachychirothere
more than a Grallator.
Newarkian late Tuvalian (Late Carnian) to late Norian age track assemblages
are well known and could even be called stereotypical. They differ substantially
from Middle Triassic age footprint assemblages described by Demathieu (1970),
Demathieu and Gand (1972), and Demathieu and Weidmann (1981) in having
abundant unquestioned dinosaurian tracks and a broader range of quadrupedal
ichnites. The middle Pekin faunule is intermediate in age and composition
between the much better known Middle Triassic and late Triassic assemblages,
and is thus critical to an understanding of the origin of dinosaur-dominated
communities of the later Mesozoic. For this reason, a concerted effort
should be made to collect more and better material from this key interval,
especially from active quarries.
ACKNOWLEDGMENTS
We are extremely grateful to the owners of both the Pomona Terra-cotta
and Borden Clay Products quarries for access to their sites for over twenty
years. We also thank Mr. James L. Mashburn for information on Pomona B
skeletal remains and for permission to photograph his private collection.
We acknowledge Hans Sues for help with comparisons to Europe. Amy Litt,
Gustav Paulay, Robert Salvia, Richard Upright, and Allen Wells are thanked
for help in the fieldwork from 1971 to 1977. I am grateful for a review
of the manuscript by Annika K. Johansson which substantially improved it.
Fieldwork during the 1970's for this paper was supported by funds supplied
through Keith S. Thomson and John H. Ostrom then of Yale University whom
PEO warmly thanks. This paper was supported by National Science Foundation
grant ATM 93-17227 and funds from Lamont-Doherty Earth Observatory. This
is Lamont-Doherty Earth Observatory contribution #5853.
REFERENCES
Axsmith, Brian J., Taylor, T. N., Delevoryas, T., Hope, R. C., 1995,
A new species of Eoginkgoites from the Upper Triassic of North Carolina,
USA: Review of Palaeobotany and Palynology, v. 85, p.189-198.
Baird, D. 1954, Chirotherium lulli, a pseudosuchian reptile from
New Jersey. Bulletin of the Museum of Comparative Zoology: v. 111, p. 115-192.
Baird, D., 1957, Triassic reptile footprint faunules from Milford, New
Jersey. Museum of Comparative Zoology Bulletin: v. 117, p. 449-520.
Baird, D. and Patterson, O. F., III, 1968, Dicynodont-archosaur fauna
in the Pekin Formation (Upper Triassic) of North Carolina [abs.] Geological
Society of America Special Paper 115, p. 11.
Biron, P. E. and Dutuit, J-M., 1981, Figurations sédimentaires
et traces d'activité au sol dans le Trias de la formation d'Argana
et de l'Ourika (Maroc). Bulletin du Museum National d'Histoire Naturelle.
Section C: Sciences de la Terre: Paleontologie, Geologie, Mineralogie.
ser. 3, no. 4, p. 399-427.
Chatterjee, S., 1985, Postosuchus, a new thecodontian reptile
from the Triassic of Texas and the origin of tyrannosaurs. Philosophical
Transactions of the Royal Society of London Ser. B: v. 309, p. 395-460.
Cornet B., 1977, The palynostratigraphy and age of the Newark Supergroup
[Ph.D. thesis]: The Pennsylvania State University, State College, 504 p.
Courel, L., Demathieu, G., Buffard, R., 1968, Empreintes de pas de vertebres
et stratigraphie du trias: Bulletin de la Société Géologique
de France, Ser. 7, v. 10, p. 275-281.
Cousminer H. L, Manspeizer W., 1976, Triassic pollen date High Atlas
and the incipient rifting of Pangea as middle Carnian: Science, v. 191,
p. 943-945.
Delevoryas, T. and Hope, R. C., 1975, Voltzia andrewsii, n. sp.,
an upper Triassic seed cone from North Carolina, U.S.A.: Review of Palaeobotany
and Palynology, v.20, p.67-74.
Demathieu, G. and Gand, G., 1972, Les Pistes dinosauroides du Trias
moyen du plateau d'Antully et leur signification paleozoologique: Bulletin
Trimestriel de la Societe d'Histoire Naturelle et des Amis du Museum d'Autun,
v.62, p.4-18.
Demathieu, G. R. and Weidmann, M., 1981, Les empreintes de pas de reptiles
dans le Trias du Vieux Emosson (Finhaut, Valais, Suisse): Eclogae Geologicae
Helvetiae, v.75, p.721-757.
Farlow, J., 1981, Estimates of dinosaur speeds from a new trackway site
in Texas: Nature, v. 294, p. 747-748.
Farlow, J. O. and R. E. Chapman. 1997. The scientific study of dinosaur
footprints; in Farlow, J. O. and Brett-Surman, M. K., eds., The
Complete Dinosaur: Indiana University Press, in press.
Fraser, N. C., and Olsen, P. E., 1996, A new dinosauromorph ichnogenus
from the Triassic of Virginia.: Jeffersoniana, no. 7, p. 1-17.
Gore, P. J. W., 1986, Depositional Framework of a Triassic Rift basin;
The Durham and Sanford subbasins of the Deep River basin, North Carolina,
in
Textoris, D. (ed.), SEPM Field Guidebook, Southeastern United States, 3rd
Annual Midyear Meeting: Raleigh, North Carolina, p. 55-115.
Haubold, H., 1971, Ichnia Amphibiorum et Reptilorum fossilium,
in
Kuhn, O., ed., Handbook of Paleoherpetology: v. 18, Fischer, Stuttgart,
124 p.
Haubold, H., 1986, Archosaur footprints at the terrestrial Triassic-Jurassic
transition: in Padian, ed., The Beginning of the Age of Dinosaurs,
Faunal Change Across the Triassic-Jurassic Boundary: Cambridge University
Press, New York, p. 189-201.
Hitchcock, E. H. 1836. Ornithichnology ? Description of the footmarks
of birds, (Ornithichnites) on New Red Sandstone in Massachusetts: American
Journal of Science 29(ser. 1), p. 307-340.
Hope, R. C. and Patterson, O. F., III, 1969, Triassic flora from the
Deep River basin, North Carolina: North Carolina Division of Mineral Resources
Special Publication 2, 22 p.
Huber, P., Lucas, S. G., and Hunt, A. P., 1993, Revised age and correlation
of the Upper Triassic Chatham Group (Deep River Basin, Newark Supergroup),
North Carolina: Southeastern Geology, v.33, p.171-193.
King, M. J. and Benton, M. J., 1996, Dinosaurs in the Early and Mid
Triassic? The footprint evidence from Britain: Palaeogeography, Palaeoclimatology,
Palaeoecology, v.122, p.213-225.
Lockley, M., 1991, Tracking Dinosaurs: A New Look at an Ancient World:
Cambridge University Press, Cambridge, 238 p.
Lucas, S. G., Hunt, A. P., and Long, R. A., 1992, The oldest dinosaurs:
Naturwissenschaften, v. 79, p. 171-172.
Lucas, S. G. and Huber, P., 1997, Vertebrate biostratigraphy and biochronology
of the non-marine Late Triassic. in LeTourneau, P. M. and Olsen,
P. E., eds., Aspects of Triassic-Jurassic Rift Basin Geoscience: Columbia
University Press, New York, in press.
Lull, R. S., 1953, Triassic life of the Connecticut Valley: State of
Connecticut, State Geological and Natural History Survey Bulletin, v. 81,
p. 1-336.
Novas, F. E., 1993, New information on the systematics and postcranial
skeleton of Herrerasaurus ischigualensis (Theropoda: Herrerasauridae) from
the Ischigualasto Formation (Upper Triassic) of Argentina: Journal of Vertebrate
Paleontology, v. 13, p. 400-423.
Olsen, P. E., 1997, Stratigraphic record of the early Mesozoic breakup
of Pangea in the Laurasia-Gondwana rift system: Annual Reviews of Earth
and Planetary Science, v. 25, p. 337-401.
Olsen, P. E., 1988, Paleoecology and paleoenvironments of the continental
early Mesozoic Newark Supergroup of eastern North America,
in Manspeizer,
W., ed., Triassic-Jurassic Rifting and the opening of the Atlantic Ocean:
Elsevier, Amsterdam, p. 185-230.
Olsen, P. E. and Baird, D., 1986, The ichnogenus
Atreipus and
its significance for Triassic Biostratigraphy: in Padian, K., ed.,
The Beginning of the Age of Dinosaurs, Faunal Change Across the Triassic-Jurassic
Boundary: Cambridge University Press, New York, p. 61-87.
Olsen, P. E. and Flynn, J., 1989, Field guide to the vertebrate paleontology
of Late Triassic rocks in the southwestern Newark Basin (Newark Supergroup,
New Jersey and Pennsylvania): The Mosasaur, v. 4, p. 1-35.
Olsen, P. E. and Galton, P.M., 1977, Triassic-Jurassic tetrapod extinctions:
are they real? Science, v. 197, p. 983-986.
Olsen, P. E. and Huber, P., 1996, The oldest Late Triassic footprint
assemblage from North America (Pekin Formation, Deep
River basin, North Carolina, USA). Southestern Geology, v. 38, no.
2, p. 77-90.
Olsen, P. E., Smith, J. B., and McDonald, N. G., 1997, Type material
of the classic theropod footprint genera Eubrontes, Anchisauripus,
and Grallator (Early Jurassic, Hartford and Deerfield basins, Massachusetts,
USA): Journal of Vertebrate Paleontology, in review.
Olsen, P. E., Schlische, R. W., and Gore, P. J. W., 1989, Field Guide
to the Tectonics, stratigraphy, sedimentology, and paleontology of the
Newark Supergroup, eastern North America: International Geological Congress,
Guidebooks for Field Trips T351, 174 p.
Padian, K. and May, C. L., 1993, The earliest dinosaurs,
in Lucas,
S. G. and Morales, M., eds., The nonmarine Triassic: Bulletin of the New
Mexico Museum of Natural History and Science, v.3, p.379-381.
Parrish, J. M., 1986, Structure and function of the tarsus in the phytosaurs
(Reptilia: Archosauria). in Padian, K., ed., The Beginning of the
Age of Dinosaurs, Faunal Change Across the Triassic-Jurassic Boundary:
Cambridge University Press, New York, p. 35-43.Reinemund, J. A., 1955,
Geology of the Deep River Coal Field North Carolina: U.S. Geological Survey
Professional Paper 246, 159 p.
Shaler, N. S. and Woodworth, J. B., 1899, Part 2; Geology of the Richmond
Basin, Virginia: U. S. Geological Survey Annual Report, v.19, Part 2, p.385-515.
Weems, R. E. and Olsen, P. E., 1997, Synthesis and revision of groups
within the Newark Supergroup, eastern North America: Geological Society
of America Bulletin, v. 109, p. 195-209.